



A hipertensão arterial é um problema de Saúde Pública, afeta 25% da população adulta mundial. Fatores genéticos e ambientais contribuem para a sua patogénese. O polimorfismo C825T da subunidade β3da Proteína G (rs5443) favorece a produção de uma variante alternativa, truncada, que facilita a sinalização intracelular, pode interferir na regulação da pressão arterial. Essa variante genética tem sido descrita como um fator de risco para a hipertensão arterial, com resultados controversos.
ObjetivoAvaliar a associação do polimorfismo C825T do gene GNβ3 com o aparecimento de hipertensão arterial, numa população portuguesa do Arquipélago da Madeira.
MétodosCom uma amostra de 1641 indivíduos (média de 50,6 ± 8,1 anos), fizemos um estudo caso‐controlo com 848 indivíduos com hipertensão arterial e 793 controlos, ajustados para o sexo e a idade. Todos os participantes colheram sangue para análises bioquímicas e foram genotipados para o polimorfismo C825T. Foi feita uma regressão logística para ver quais as variáveis que se relacionam com a hipertensão arterial. A análise dos dados foi feita com o software estatístico SPSS, versão 19.0. Usamos como limiar de significância o valor de p < 0,05.
ResultadosEncontramos uma associação significativa entre o polimorfismo C825T e o aparecimento de hipertensão arterial (odds ratio = 1,275; IC 95% (1,042–1,559); p = 0,018) no modelo dominante, após análise multivariada.
ConclusãoO polimorfismo C825T da subnidade β3da Proteína G está associado, de forma significativa e independente, com o aparecimento hipertensão arterial na nossa população.
Hypertension is an important public health problem, affecting about 25% of the adult population worldwide.1 Genetic and environmental factors contribute to its pathogenesis. The T allele of the C825T polymorphism of the beta 3 subunit of G protein (rs5443) leads to the production of a truncated variant that enhances intracellular signaling and may interfere with the regulation of blood pressure. This genetic variant has been described as a risk factor for hypertension, although study results are controversial.
ObjectiveThe objective of this study was to analyze the association of the C825T polymorphism of the GNB3 gene with the occurrence of hypertension in a Portuguese population from the Madeira archipelago.
MethodsA case‐control study was performed with 1641 Caucasian individuals (mean age 50.6±8.1 years), 848 with hypertension and 793 controls. Blood was collected from all participants for biochemical and genetic analysis, including genotyping of the C825T polymorphism. Logistic regression analysis was performed to determine which variables were significantly associated with the onset of hypertension. Statistical analyses were performed using IBM SPSS version 19.0 and p‐values <0.05 were considered statistically significant.
ResultsIn our study, there was a significant association between the C825T polymorphism of the GNB3 gene and the occurrence of hypertension (odds ratio 1.275; 95% confidence interval 1.042‐1.559; p=0.018) in the dominant model, after multivariate analysis.
ConclusionWe conclude that the C825T polymorphism of the beta 3 subunit of G protein is significantly and independently associated with the occurrence of hypertension in the study population.
A hipertensão arterial (HTA) é um problema importante de saúde pública, afeta cerca de 25% da população adulta mundial1. É um fator de risco de doença cardiovascular e de progressão para insuficiência renal, independentemente de outros fatores de risco vascular2,3. Na população adulta portuguesa, a sua prevalência global é de 42% (44,4% nos homens e 40,2% nas mulheres)4.
Nas últimas décadas, muitos estudos têm sido feitos no sentido de esclarecer qual o contributo genético no desenvolvimento da HTA. Sabe‐se que de 20‐40% da variação da pressão arterial é determinada geneticamente5,6. No entanto, a genética molecular da HTA ainda permanece pouco esclarecida.
As proteínas G e seus recetores têm um papel crucial na sobrevivência celular. Essas proteínas fazem parte de uma superfamília de proteínas que, no estado inativo, encontram‐se acopladas a recetores da membrana celular. Quando ativadas, por estímulos adequados, graças a propriedades funcionais e estruturais, migram pelo citosol e ativam múltiplos efetores, como sejam enzimas, canais iónicos, hormonas, neurotransmissores e fatores autócrinos e parácrinos7, consumam a transmissão de sinais. Esse é um processo de ativação dos eventos intracelulares por estímulos externos.
A transdução de estímulos externos em sinais intracelulares é comprovadamente um dos fatores de grande importância para a perpetuação das características viáveis na escala evolutiva. Tendo em vista o seu papel abrangente, a deficiência da expressão ou formas alteradas dessas importantes proteínas podem levar a distúrbios metabólicos globais ou restritos8.
As proteínas G têm uma estrutura heterotrimérica, composta por três subunidades denominadas alfa (α), beta (β) e gama (γ), codificadas por genes distintos. A subunidade alfa (α) é a mais característica de cada proteína G e é essa subunidade que interage com o recetor, liga‐se ao GTP e regula os sistemas efetores9. As subunidades β e γ estão associadas por ligações não covalentes e, quando se encontram ligadas à subunidade α, configuram o estado inativo da Proteína G10. Os dímeros β e γ, para além de servir para ancorar a subunidade α, também têm influência em processos celulares específicos11,12 e podem modular a atividade de determinados efetores13. Devido ao seu papel crucial na função de muitos tipos de células, as anormalidades genéticas nas subunidades das proteínas G têm o potencial de estar envolvidas na etiologia de uma vasta gama de situações clínicas.
O gene que codifica a subunidade β3da Proteína G está localizado no cromossoma 12p13. O polimorfismo da subunidade β3da Proteína G (C825T) rs5443 situa‐se no exão 10 do gene GNβ3, onde há uma substituição da citosina (C) pela timidina (T) no nucleótido 825. O alelo 825T está associado com a ocorrência dum splicing alternativo dentro do exão 9, no qual os nucleótidos 498‐620 são eliminados14,15. Como resultado, os aminoácidos 167 a 197 estão ausentes na proteína codificada (designada Gβ3‐s). Essa variante truncada plice (Gβ3‐s) é uma proteína funcional que confere uma ativação aumentada das proteínas G, facilita a sinalização intracelular14,16. Dado que a ativação da proteína G é o evento‐chave na transdução de sinal intracelular, o polimorfismo C825T tem um papel importante na etiologia duma vasta gama de situações clínicas, bem como na resposta a fármacos17. Essa variante genética foi descrita como associada a várias patologias, HTA14,15,18–20, obesidade21, depressão22 e doenças cardiovasculares23. Além disso, o alelo 825T pode servir como marcador farmacogenético na previsão de respostas a vários fármacos como sejam: diuréticos, antidepressivos, sildenafil, clonidina, ngiotensina II e endotelina‐117.
No entanto, os resultados de alguns estudos não demonstraram associação dessa variante genética com a HTA24‐26. Há estudos que confirmam e outros que rejeitam a ligação do polimorfismo da subunidade β3da Proteína G (C825T) à HTA, justifica‐se avaliar a sua eventual associação numa população portuguesa.
ObjetivoAvaliar a eventual associação do polimorfismo C825T da Proteína G (GNβ3, rs5443) com o aparecimento de HTA, numa população portuguesa do Arquipélago da Madeira.
MétodosProtocolo do estudo e aprovação éticaEste estudo foi aprovado pela Comissão de Ética do Sesaram (Serviço de Saúde da Região Autónoma da Madeira) e foi conduzido de acordo com as diretrizes éticas internacionais da Declaração de Helsínquia de 2013.
Todos os indivíduos assinaram o termo de consentimento informado e foram informados, pelo médico internista com mais de 10 anos de experiência, acerca da colheita de sangue para análise e armazenamento e reutilização de ADN e de dados clínicos relevantes. As amostras de sangue para fins genéticos são mantidas no nosso biobanco de pesquisa hospitalar de acordo com os artigos 6, 15 e 16da Declaração Universal da Unesco sobre Bioética e Direitos Humanos.
População do estudoEste estudo foi feito no Arquipélago da Madeira, Portugal, cuja população é de cerca de 300 000 habitantes. Os indivíduos participantes no estudo são de etnia branca, naturais do Arquipélago da Madeira, e os seus progenitores (1.ª e 2.ª geração) são também nascidos nesse arquipélago e da mesma etnia. Procuramos dessa forma que essa população de indivíduos fosse geneticamente homogénea e representasse assim uma população da Europa do Sul.
Com uma amostra de 1641 indivíduos com média 50,6 ± 8,1 anos, 848 (49,9%) eram do género masculino, selecionados sequencialmente das consultas de medicina geral e familiar e de medicina interna do Hospital Central do Funchal.
Constituímos dois grupos, com a nossa amostra, consoante tinham ou não HTA, procuramos que não houvesse diferenças significativas entre ambos os grupos em termos de sexo e idade.
Hipertensão arterialA HTA foi considerada se os indivíduos à entrada no estudo já estivessem diagnosticados e/ou cumprissem medicação anti‐hipertensiva havia mais de três meses ou ainda se fossem hipertensos recém‐diagnosticados com pressão arterial sistólica (PAS) e/ou pressão arterial diastólica (PAD) ≥ 140/90mmHg, avaliada em, pelo menos, três ocasiões27.
Foram excluídos todos os indivíduos com hipertensão secundária, tais como hiperaldosteronismo primário ou doença renal clinicamente relevante, e os que receberam medicação para outras indicações que pode afetar a pressão arterial.
Os controlos normotensos nunca tinham feito medicação anti‐hipertensiva e apresentavam PAS/ PAD < 140/90mmHg.
Foi avaliada a pressão arterial a todos os indivíduos, após 10min de repouso, em posição sentada, no braço direito, com o uso de um esfigmomanómetro Welch Allyn padrão (fases I a V), e foi registada a média de três leituras feitas com intervalo de dois min27.
Recolha de dadosTodos os participantes responderam a um questionário padronizado em relação a idade, sexo, estilo de vida, tabagismo, consumo de álcool, história pessoal e familiar de eventos cardiovasculares, nomeadamente acidente vascular cerebral e doença das artérias coronárias e história familiar de HTA.
Foram avaliados, em todos os participantes, os parâmetros antropométricos. Foi medida a altura em centímetros e o peso em quilogramas e calculado o Índice de Massa Corporal (IMC) com o uso da fórmula (kg / m2)28. O IMC foi dividido em três subgrupos: normal (< 25kg/m2), excesso de peso (25 ≤ IMC < 30kg/m2) e obesidade (IMC ≥ 30kg/m2)28. O perímetro abdominal foi avaliado e considerado elevado quando superior a 94cm para o sexo masculino e a 80cm para o feminino29. A razão cintura/anca foi calculada, foi considerada elevada para um valor > 0,9 para o sexo masculino e > 0,8 para o feminino29.
A velocidade de onda de pulso (VOP) foi determinada por tonometria através do Complior como previamente descrito30. Um valor de VOP > 10 m/s2 foi considerado elevado. A diabetes mellitus (DM) foi definida por um nível de glicose em jejum ≥126mg/dl e/ou um nível de glicose ≥ 200mg/dl e/ou uma história de tratamento para DM31.
Sedentarismo foi definido quando os indivíduos não praticavam pelo menos 150 minutos/semana de atividade física moderada a vigorosa29.
Consideraram‐se fumadores aqueles que à data da entrada no estudo tinham hábitos tabágicos e dividiu‐se essa variável em três subgrupos, consoante o número de cigarros consumidos por dia: de um a 10 cigarros; entre 10 a 20 cigarros e mais de 20 cigarros. Os indivíduos que ingeriam álcool à entrada do estudo foram considerados como tendo hábitos alcoólicos. Quantificou‐se o consumo de álcool em gramas por semana, foi considerado excesso quando superior a 70 gr/semana no sexo masculino e 50 gr/semana no feminino.
Análise bioquímicaAs amostras de sangue foram extraídas após 14‐16 horas de jejum e o plasma foi preparado para a quantificação dos perfis biológicos. As análises bioquímicas foram feitas no Laboratório Central do Hospital, de acordo com as técnicas usuais.
Para determinar o colesterol total, HDL, LDL, triglicéridos e glucose, as amostras de sangue foram colocadas em tubos secos, centrifugadas durante meia hora a 3500g e posteriormente quantificadas por uma técnica enzimática com o uso de um analisador automático «AU 5400» (Beckman Coulter). Os marcadores bioquímicos de risco lipoproteína (a), apoliproteína B e a Proteína C‐reativa de alta sensibilidade (PCR‐as) foram quantificados por um aparelho automático «AU 5400» (Beckman Coulter) pela técnica de imunoturbidimetria. Em relação ao fibrinogénio, a colheita também foi feita com o doente em jejum, para um tubo com citrato de sódio, com o uso de um analisador automático «ACL TOP 700».
Análise genética do polimorfismo C825T do gene GNβ3O ADN genómico foi extraído dos leucócitos do sangue periférico com o uso do método de salting‐out. A análise genotípica foi efetuada com sondas oligonucleotídicas marcadas com fluorescência específica para cada um dos alelos num ensaio que combina a técnica convencionada de PCR e a técnica TaqMan (Applied Biosystems). Os primers e as sondas foram as pré‐estabelecidas pelo fornecedor (TaqMan SNP Genotyping Assays, Applied Biosystems) para o polimorfismo C825T do gene GNβ3 (rs5443), os oligonucleótidos foram sintetizados e os marcadores fluorogénicos FAM e VIC acoplados às extremidades 5’ das sondas de modo a alcançar a discriminação alélica. A reação de polimerização em dois passos consistia em 40 ciclos de desnaturação a 92°C durante 15 segundos e de primer annealing e extensão a 60°C durante um minuto e foi efetuada num aparelho 7.300 Real‐Time PCR System (Applied Biosystems). Os genótipos foram determinados com o 7.300 System SDS Software (Applied Biosystems) sem qualquer conhecimento prévio dos dados clínicos individuais (Fig. 1).
Análise estatística.")
As variáveis categóricas são apresentadas pela frequência e a sua respetiva percentagem, com o uso do teste de qui‐quadrado. As variáveis contínuas são expressas com a média ± desvio padrão ou mediana (mínimo–máximo). Para comparar as variáveis contínuas foram usados os testes t de Student ou o teste não paramétrico de Mann‐Whitney, conforme apropriado.
O equilíbrio de Hardy‐Weinberg foi calculado para cada um dos genes com o uso do teste de qui‐quadrado. Um valor de p inferior a 5% foi considerado estatisticamente significativo, enquanto o valor Pb (p dividido pelo número total de comparações) foi considerado após a correção de Bonferroni.
Para determinar, por análise multivariada, quais as variáveis associadas ao aparecimento de HTA, foi elaborada uma regressão logística com o uso do método Forward Wald.
A força de associação foi determinada pela odds ratio (OR) e pelos respetivos intervalos de confiança (IC) a 95%.
Os dados estatísticos foram analisados com o uso do software estatístico SPSS (Statistical Package for the Social Sciences) versão 19.0. Usamos como limiar de significância o valor de p < 0,05.
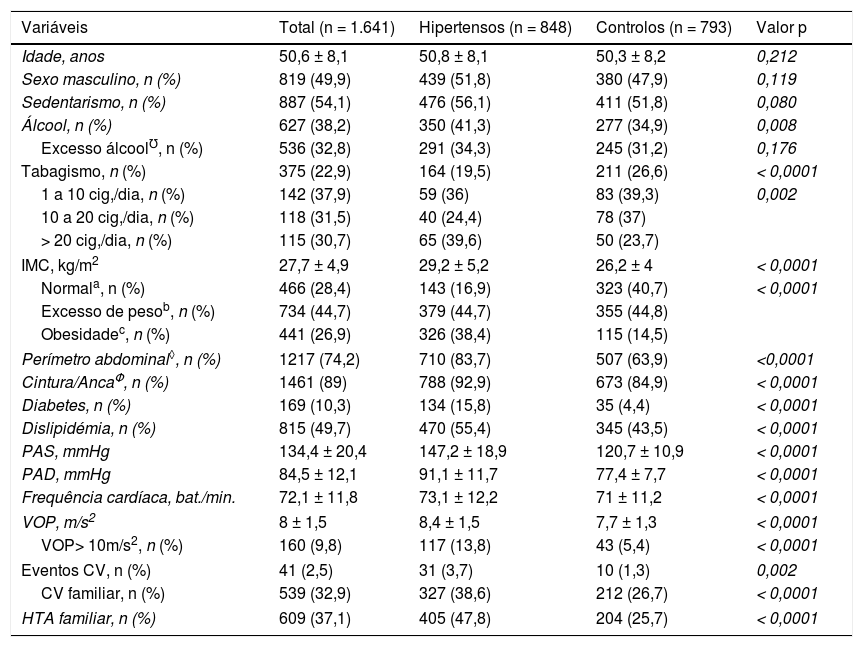
ResultadosCaracterísticas dos participantes do estudoApresentamos as características basais dos participantes do estudo na Tabela 1. A população estudada é composta por 1641 indivíduos, 848 casos com HTA (idade média de 50,8 ± 8,1; 51,8% do sexo masculino) e 793 controlos sem HTA (idade média de 50,3 ± 8,2; 47,9% do sexo masculino). As variáveis género e idade foram ajustadas entre os grupos, não houve diferenças significativas (Tabela 1).
Características demográficas e clínicas dos participantes do estudo
| Variáveis | Total (n = 1.641) | Hipertensos (n = 848) | Controlos (n = 793) | Valor p |
|---|---|---|---|---|
| Idade, anos | 50,6 ± 8,1 | 50,8 ± 8,1 | 50,3 ± 8,2 | 0,212 |
| Sexo masculino, n (%) | 819 (49,9) | 439 (51,8) | 380 (47,9) | 0,119 |
| Sedentarismo, n (%) | 887 (54,1) | 476 (56,1) | 411 (51,8) | 0,080 |
| Álcool, n (%) | 627 (38,2) | 350 (41,3) | 277 (34,9) | 0,008 |
| Excesso álcool℧, n (%) | 536 (32,8) | 291 (34,3) | 245 (31,2) | 0,176 |
| Tabagismo, n (%) | 375 (22,9) | 164 (19,5) | 211 (26,6) | < 0,0001 |
| 1 a 10 cig,/dia, n (%) | 142 (37,9) | 59 (36) | 83 (39,3) | 0,002 |
| 10 a 20 cig,/dia, n (%) | 118 (31,5) | 40 (24,4) | 78 (37) | |
| > 20 cig,/dia, n (%) | 115 (30,7) | 65 (39,6) | 50 (23,7) | |
| IMC, kg/m2 | 27,7 ± 4,9 | 29,2 ± 5,2 | 26,2 ± 4 | < 0,0001 |
| Normala, n (%) | 466 (28,4) | 143 (16,9) | 323 (40,7) | < 0,0001 |
| Excesso de pesob, n (%) | 734 (44,7) | 379 (44,7) | 355 (44,8) | |
| Obesidadec, n (%) | 441 (26,9) | 326 (38,4) | 115 (14,5) | |
| Perímetro abdominal◊, n (%) | 1217 (74,2) | 710 (83,7) | 507 (63,9) | <0,0001 |
| Cintura/AncaΦ, n (%) | 1461 (89) | 788 (92,9) | 673 (84,9) | < 0,0001 |
| Diabetes, n (%) | 169 (10,3) | 134 (15,8) | 35 (4,4) | < 0,0001 |
| Dislipidémia, n (%) | 815 (49,7) | 470 (55,4) | 345 (43,5) | < 0,0001 |
| PAS, mmHg | 134,4 ± 20,4 | 147,2 ± 18,9 | 120,7 ± 10,9 | < 0,0001 |
| PAD, mmHg | 84,5 ± 12,1 | 91,1 ± 11,7 | 77,4 ± 7,7 | < 0,0001 |
| Frequência cardíaca, bat./min. | 72,1 ± 11,8 | 73,1 ± 12,2 | 71 ± 11,2 | < 0,0001 |
| VOP, m/s2 | 8 ± 1,5 | 8,4 ± 1,5 | 7,7 ± 1,3 | < 0,0001 |
| VOP> 10m/s2, n (%) | 160 (9,8) | 117 (13,8) | 43 (5,4) | < 0,0001 |
| Eventos CV, n (%) | 41 (2,5) | 31 (3,7) | 10 (1,3) | 0,002 |
| CV familiar, n (%) | 539 (32,9) | 327 (38,6) | 212 (26,7) | < 0,0001 |
| HTA familiar, n (%) | 609 (37,1) | 405 (47,8) | 204 (25,7) | < 0,0001 |
Cig.: cigarros; CV: cardiovasculares; HTA: hipertensão arterial; IMC: índice de massa corporal; PAD: pressão arterial diastólica; PAS: pressão arterial sistólica; ℧ > 70gr/dia para os homens e > 50gr/dia para as mulheres; VOP: Velocidade de onda de pulso
Verifica‐se que houve diferença significativa em relação às seguintes variáveis: consumo de álcool (p = 0,008), IMC e obesidade (p < 0,0001), perímetro abdominal (p < 0,0001) e cintura/anca (p < 0,0001), apresentaram valores médios mais elevados no grupo dos hipertensos em relação aos controlos. Da mesma maneira, a DM, dislipidémia, pressão arterial sistólica e distólica, frequência cardíaca e a VOP foram mais frequentes no grupo dos indivíduos hipertensos em relação aos normotensos, com significância estatística (p < 0,0001). No grupo dos hipertensos, havia mais indivíduos com história pessoal (p = 0,002) e familiar (p < 0,0001) de eventos cardiovasculares, assim como história familiar de HTA (p < 0,0001). Constatou‐se também que havia uma maior percentagem de fumadores no grupo dos controlos em todos os subgrupos, exceto nos que fumavam mais de 20 cigarros/dia, em relação ao grupo dos hipertensos e com significância estatística (p < 0,0001).
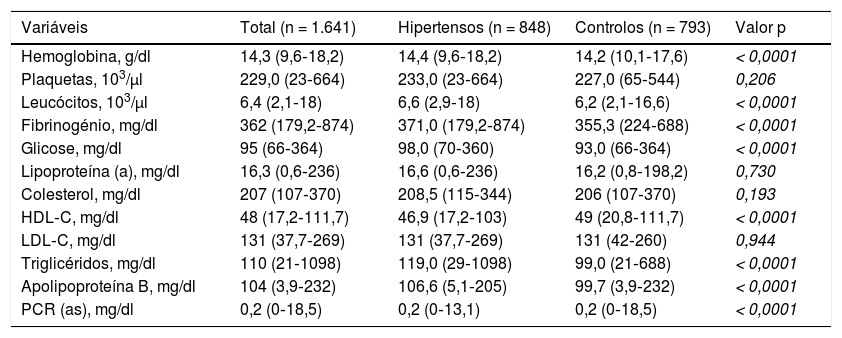
Em relação às variáveis bioquímicas (Tabela 2), houve diferença significativa entre os dois grupos em relação à hemoglobina (p < 0,0001), leucócitos (p < 0,0001), fibrinogénio (p < 0,0001), HDL‐colesterol (p < 0,0001), triglicerídeos (p < 0,0001), apolipoproteína B (p < 0,0001), glicose em jejum (p < 0,0001) e PCR (as) (p < 0,0001), que também apresentaram valores médios mais elevados no grupo dos hipertensos em relação aos controlos. Em relação às variáveis colesterol total, LDL‐ colesterol, lipoproteína (a) e plaquetas não houve diferença significativa entre os dois grupos (p > 0,05).
Características bioquímicas dos participantes do estudo
| Variáveis | Total (n = 1.641) | Hipertensos (n = 848) | Controlos (n = 793) | Valor p |
|---|---|---|---|---|
| Hemoglobina, g/dl | 14,3 (9,6‐18,2) | 14,4 (9,6‐18,2) | 14,2 (10,1‐17,6) | < 0,0001 |
| Plaquetas, 103/μl | 229,0 (23‐664) | 233,0 (23‐664) | 227,0 (65‐544) | 0,206 |
| Leucócitos, 103/μl | 6,4 (2,1‐18) | 6,6 (2,9‐18) | 6,2 (2,1‐16,6) | < 0,0001 |
| Fibrinogénio, mg/dl | 362 (179,2‐874) | 371,0 (179,2‐874) | 355,3 (224‐688) | < 0,0001 |
| Glicose, mg/dl | 95 (66‐364) | 98,0 (70‐360) | 93,0 (66‐364) | < 0,0001 |
| Lipoproteína (a), mg/dl | 16,3 (0,6‐236) | 16,6 (0,6‐236) | 16,2 (0,8‐198,2) | 0,730 |
| Colesterol, mg/dl | 207 (107‐370) | 208,5 (115‐344) | 206 (107‐370) | 0,193 |
| HDL‐C, mg/dl | 48 (17,2‐111,7) | 46,9 (17,2‐103) | 49 (20,8‐111,7) | < 0,0001 |
| LDL‐C, mg/dl | 131 (37,7‐269) | 131 (37,7‐269) | 131 (42‐260) | 0,944 |
| Triglicéridos, mg/dl | 110 (21‐1098) | 119,0 (29‐1098) | 99,0 (21‐688) | < 0,0001 |
| Apolipoproteína B, mg/dl | 104 (3,9‐232) | 106,6 (5,1‐205) | 99,7 (3,9‐232) | < 0,0001 |
| PCR (as), mg/dl | 0,2 (0‐18,5) | 0,2 (0‐13,1) | 0,2 (0‐18,5) | < 0,0001 |
As variáveis bioquímicas são apresentadas pela mediana (mínimo ‐ máximo).
HDL‐C: colesterol das lipoproteínas de alta densidade; LDL‐C: colesterol das lipoproteínas de baixa densidade; PCR (as): proteína C reativa de alta sensibilidade; significância estatística para p < 0,05.
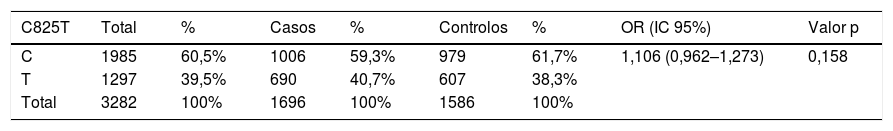
Apresentamos as frequências dos alelos do polimorfismo genético C825T do gene GNβ3 na Tabela 3. A frequência dos alelos C e T do polimorfismo C825T na população total da nossa amostra foi de 60,5% e de 39,5%, respetivamente. No grupo dos casos e controlos seguiu a mesma tendência, atingiu um valor de OR de 1,106, mas sem significância estatística (p = 0,158) (Tabela 3).
Frequências alélicas do polimorfismo genético C825T do gene GNβ3
| C825T | Total | % | Casos | % | Controlos | % | OR (IC 95%) | Valor p |
|---|---|---|---|---|---|---|---|---|
| C | 1985 | 60,5% | 1006 | 59,3% | 979 | 61,7% | 1,106 (0,962–1,273) | 0,158 |
| T | 1297 | 39,5% | 690 | 40,7% | 607 | 38,3% | ||
| Total | 3282 | 100% | 1696 | 100% | 1586 | 100% |
IC: Intervalo de confiança; OR: odds ratio; significância estatística para p < 0,05.
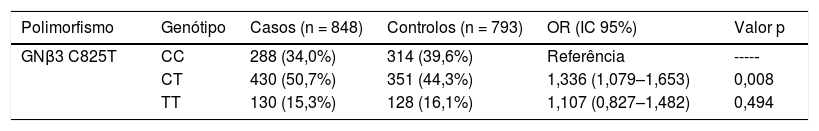
A frequência de cada genótipo em ambos os grupos, casos e controlos, foi encontrada em conformidade com o equilíbrio de Hardy‐Weinberg (Tabela 4). O genótipo mais frequente, no modelo aditivo, foi o heterozigoto CT, que apresentou valores de 50,7% e 44,3% nos casos e controlos, respetivamente, quando comparado com o genótipo homozigoto de referência CC. Esse genótipo acarretou um risco maior de 1,336 (p = 0,008) em relação ao homozigoto mutado (OR = 1,107; p = 0,494) (tabela 4).
Genótipos do polimorfismo genético C825T do gene GNβ3 e risco de hipertensão arterial
| Polimorfismo | Genótipo | Casos (n = 848) | Controlos (n = 793) | OR (IC 95%) | Valor p |
|---|---|---|---|---|---|
| GNβ3 C825T | CC | 288 (34,0%) | 314 (39,6%) | Referência | ----‐ |
| CT | 430 (50,7%) | 351 (44,3%) | 1,336 (1,079–1,653) | 0,008 | |
| TT | 130 (15,3%) | 128 (16,1%) | 1,107 (0,827–1,482) | 0,494 |
IC: intervalo de confiança; OR: odds ratio; significância estatística para p < 0,05.
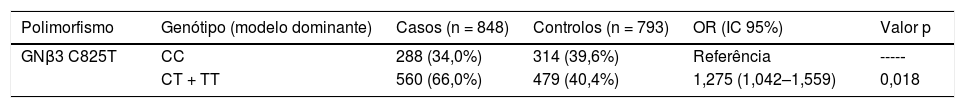
No modelo dominante (Tabela 5), os indivíduos com genótipo (CT ou TT) tinham um maior risco de desenvolver HTA (OR = 1,275; IC 95% = 1,042–1,559) quando comparados com o genótipo de referência (CC) (p = 0,018).
Genótipos do polimorfismo genético C825T do gene GNβ3 e risco de hipertensão arterial (modelo dominante)
| Polimorfismo | Genótipo (modelo dominante) | Casos (n = 848) | Controlos (n = 793) | OR (IC 95%) | Valor p |
|---|---|---|---|---|---|
| GNβ3 C825T | CC | 288 (34,0%) | 314 (39,6%) | Referência | ----‐ |
| CT + TT | 560 (66,0%) | 479 (40,4%) | 1,275 (1,042–1,559) | 0,018 |
IC: Intervalo de confiança; OR: odds ratio; significância estatística para p < 0,05.
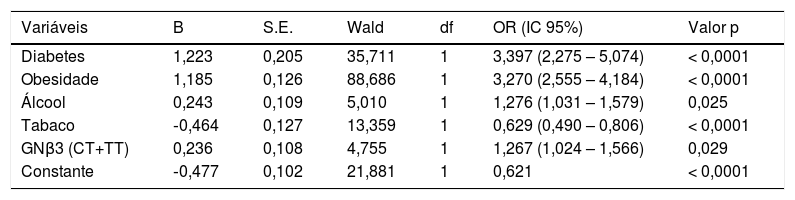
Foi feita uma análise de regressão logística (Tabela 6), as variáveis que permaneceram na equação foram: DM, obesidade, consumo de álcool e os genótipos CT e TT do polimorfismo GNβ3, associaram‐se de forma significativa e independente ao aparecimento de HTA (Tabela 6).
Análise de regressão logísticaa com o modelo genético dominante do polimorfismo GNβ3 e as variáveis de confundimento
| Variáveis | B | S.E. | Wald | df | OR (IC 95%) | Valor p |
|---|---|---|---|---|---|---|
| Diabetes | 1,223 | 0,205 | 35,711 | 1 | 3,397 (2,275 – 5,074) | < 0,0001 |
| Obesidade | 1,185 | 0,126 | 88,686 | 1 | 3,270 (2,555 – 4,184) | < 0,0001 |
| Álcool | 0,243 | 0,109 | 5,010 | 1 | 1,276 (1,031 – 1,579) | 0,025 |
| Tabaco | ‐0,464 | 0,127 | 13,359 | 1 | 0,629 (0,490 – 0,806) | < 0,0001 |
| GNβ3 (CT+TT) | 0,236 | 0,108 | 4,755 | 1 | 1,267 (1,024 – 1,566) | 0,029 |
| Constante | ‐0,477 | 0,102 | 21,881 | 1 | 0,621 | < 0,0001 |
Os hábitos tabágicos surgem na equação como variável protetora em relação ao aparecimento de HTA. O sedentarismo saiu da equação, não influenciou o aparecimento de HTA (Tabela 6).
DiscussãoNo presente estudo, investigámos a existência de uma associação entre a variante GNβ3 825T da Proteína G e a HTA, numa população portuguesa do Arquipélago da Madeira. Os indivíduos com o modelo genético dominante apresentaram um risco acrescido de desenvolver HTA (OR = 1,275; IC 95% = 1,042–1,559; p = 0,018), mesmo após a análise multivariada, com correção para os fatores de confusão (obesidade, DM, sedentarismo, ingestão de álcool e hábitos tabágicos). Esses resultados demonstram que, nessa população, as variantes da Proteína G se associam à HTA de forma independente dos fatores de risco de HTA já conhecidos, ou seja, acrescentam risco de HTA aos fatores de risco tradicionais.
Na nossa população, a frequência encontrada para o alelo T foi de 39,5% nessa população, o que é superior à relatada em outras populações de etnia branca (37,7%), é inferior à descrita para os asiáticos (48,4%) e para os negros americanos (76,3%)19.
Os presentes resultados, inovadores na população portuguesa, vão na sequência de alguns estudos com resultados semelhantes.
Pela primeira vez, Siffert et al.14 demonstraram, num estudo com 426 pacientes, uma associação significativa do polimorfismo no gene GNβ3 sensível à toxina pertússis (GNβ3) com a HTA, (OR =1,79; IC 95% = 1,05–3,05; p = 0,03) numa população da Alemanha. Posteriormente, muitos estudos em nível mundial e em diferentes grupos étnicos encontraram associação entre o polimorfismo do GNβ3 C825T com a HTA14,15,17–19 ou com a variação da pressão arterial32–34.
Benjafield et al.15, num estudo feito na etnia branca concluíram que a variante GNβ3 825T da Proteína G se associava com a HTA (OR = 2,3; IC 95% = 1,7–3,3)15.
Também numa população egípcia encontrou‐se a associação dessa variante genética com a HTA (OR= 1,4; IC 95% = 0,9–2,1) no modelo genético dominante e que aumentava significativamente no modelo recessivo (OR=2,3; IC 95% = 1,5–3,7)20.
Zheng et al19, numa meta‐análise que envolveu 36 802 indivíduos, confirmaram uma associação significativa entre o polimorfismo GNβ3 (C825T) e o risco global de HTA em brancos e chineses, mas essa associação não foi comprovada noutras populações asiáticas19.
Em contraponto, Guo et al.26, numa meta‐análise, sugeriram pela análise global das evidências que o alelo GNβ3 825T pode ser um bom indicador de risco de HTA, mas não evidenciou associação com a HTA em asiáticos e brancos26.
Vários mecanismos fisiopatológicos poderão justificar o papel importante da variante genética GNβ3 C825T rs5443, no desenvolvimento de HTA.
Siffert et al.14 descreveram que a variante truncada splice ‐ Gβ3‐s é uma proteína funcional que provoca uma atividade aumentada das proteínas G nos sistemas reconstituídos, promove melhor sinalização intracelular14.
Uma pressão arterial mais elevada deve‐se ao facto duma maior sensibilidade às hormonas pressores vasoativas, aquelas que transmitem os seus sinais através de proteínas GNβ335.
Esse conceito é demonstrado em estudos in vivo, provado pelo facto de os indivíduos portadores da variante genética GNβ3 825T terem maior reatividade vascular na estimulação de recetores α1 ‐ adrenérgicos coronários36.
Meirhaeghe et al.16 avaliaram a vasomotilidade das artérias coronárias em resposta à injeção intravenosa dum fármaco vasoconstritor (metil‐ergonovina) e concluíram que os indivíduos portadores de pelo menos um alelo T do GNβ3 apresentaram maior suscetibilidade à vasoconstrição quando comparados com os do genótipo selvagem CC. A subunidade Gβ3‐s está associada a aumento da atividade dessa proteína, pode, portanto, incrementar a atividade do α1A‐adrenoreceptor acoplado às proteínas G, explica a maior vasoconstrição observada em resposta ao fármaco vasoconstritor, em indivíduos com o alelo GNβ3 825T.
Outra hipótese é defendida por Siffert et al.14, de que o início da HTA nos portadores do alelo T do GNβ3 poderá não se dever à vasoconstrição aumentada por hormonas vasoconstritoras como a noradrenalina e a angiotensina II que ativam heterotrímeros das Proteínas G não sensíveis à toxina pertússis, mas sim resultar da proliferação das células musculares lisas vasculares que levam gradualmente à hipertrofia vascular14.
Fizemos na nossa população uma análise de regressão logística na qual foram estudadas as variáveis de confundimento, como sejam obesidade, DM, hábitos alcoólicos, tabagismo e o modelo genético dominante do polimorfismo GNβ3, a fim de avaliar quais as variáveis que se relacionavam de forma significativa e independente com o aparecimento da HTA. A obesidade e a DM são variáveis que conferiram risco significativo de aparecimento de HTA com um OR de 3,270 (IC 95% = 2,555–4,184; p < 0,0001) e 3,397 (IC 95% = 2,275–5,074; p < 0,0001), respetivamente. O polimorfismo GNβ3 também se manteve na equação, com um risco (OR) de 1,276 (IC 95% = 1,024–1,566; p = 0,029), o que demonstra que se associa de forma significativa e independente com o aparecimento de HTA.
Segundo os resultados do nosso estudo no grupo dos controlos havia uma maior percentagem de indivíduos com hábitos tabágicos do que no grupo dos hipertensos em todos os subgrupos, exceto nos fumadores de mais de 20 cigarros por dia. Após a análise multivariada, os hábitos tabágicos surgiram na equação como variável protetora em relação ao aparecimento de HTA. Isso se deve ao facto de ser considerado ter hábitos tabágicos aquando à entrada do estudo, portanto os indivíduos que eram fumadores ativos, e não aqueles que tinham tido história de hábitos tabágicos. Salienta‐se o facto de que os indivíduos hipertensos, à entrada do estudo, já estavam a ser seguidos em consultas médicas, em abordagem geral do risco cardiovascular, o que terá levado a que alguns deixassem de fumar. No entanto, no nosso estudo verifica‐se que, para os indivíduos com maior consumo tabágico, torna‐se mais difícil a evicção desse hábito. Sabemos que a obesidade é também um importante fator de risco para o desenvolvimento de HTA. Estudos transversais demonstram que obesidade está associada a níveis mais elevados de pressão arterial, de ganho de peso ao longo da vida, e é um importante preditor de desenvolvimento de HTA37–40. Vários estudos, em populações de várias etnias, demonstraram a associação do alelo T do polimorfismo GNβ3 com a obesidade41,42, o que pode afetar as variações da pressão arterial43,44.
Outros estudos que correlacionem a anormalidade dos complexos mecanismos transdutores a nível celular com outros processos patológicos podem ser a base para descobertas farmacológicas importantes para a terapêutica da HTA e de muitas outras doenças.
Pontos fortes do estudoO presente estudo é o primeiro estudo caso‐controlo feito na população da Ilha da Madeira, uma população portuguesa geneticamente homogénea45–47 e relativamente isolada, em que se investiga a associação do polimorfismo GNβ3 C825T com a suscetibilidade para a HTA.
Essa população, com as características citadas, representa uma mais‐valia para o mapeamento de distúrbios raros e, além disso, segundo vários pesquisadores, o estudo de populações culturalmente e geneticamente isoladas que apresentam um modo de vida, hábitos alimentares e ambiente natural idênticos pode reduzir a variação ambiental48.
ConclusãoNo estudo, o polimorfismo GNβ3 C825T associou‐se de forma significativa e independente com o aparecimento de HTA, numa população portuguesa (OR = 1,275; IC 95% = 1,042–1,559; p = 0,018) no modelo dominante, mesmo após análise de regressão logística, na qual foram estudadas outras variáveis que podem também associar‐se ao aparecimento de HTA.
A identificação de genes candidatos e a compreensão da sua função contribuem para o estabelecimento duma base molecular específica para HTA. Serão necessários mais estudos com tamanhos de amostra maiores, em diferentes populações, para determinar se o polimorfismo GNβ3 C825T desempenha um papel relevante na etiologia da HTA.
FinanciamentoPrograma Operacional de Valorização do Potencial Económico e Coesão Territorial da Região Autónoma da Madeira (Intervir+).
Conflitos de interesseOs autores declaram não haver conflitos de interesse.
